Memória de Longo Prazo
Estudos anteriores haviam demonstrado que, ao se estimular a via que se projeta do córtex entorrinal às células granulares do hipocampo, a resposta dessas células podia ser medida com um eletrodo de registro. Bliss e seus colegas mediram a resposta normal a um estímulo elétrico isolado e depois aplicaram uma longa série de sinais de alta freqüência - chamada de estimulação tetânica - a esta via. Após a estimulação tetânica, um estímulo elétrico isolado provocou disparo maior das células granulares que o observado anteriormente. O efeito intensificado persistiu por até três dias. Este fenômeno de LTP era precisamente o tipo de aumento da força neuronal que poderia ser capaz de manter a memória. A LTP é considerada agora um modelo para o aprendizado e a memória.
A LTP ocorre pela atividade do receptor NMDA (N-metil-D-aspartato). Esta molécula está inserida nos dendritos das células granulares, das células de CAI do hipocampo e nos neurônios de toda a extensão do neocórtex. Assim como outros receptores neuronais, o receptor NMDA é ativado por um neurotransmissor - glutamato, neste caso. O glutamato abre momentaneamente um canal não-NMDA no dendrito da célula granular, permitindo um fluxo de sódio do espaço extracelular para dentro do neurônio. Este influxo causa a despolarização da célula granular. Se a despolarização for suficiente, a célula granular dispara, transmitindo informações para outros neurônios.
Diferentemente de outros receptores neuronais, o NMDA possui uma propriedade adicional. Se uma ativação adicional do glutamato ocorrer enquanto a célula granular estiver despolarizada, um segundo canal se abre, permitindo o influxo de cálcio. Acredita-se que o cálcio aja como um segundo mensageiro, iniciando uma série de eventos intracelulares que culminam em mudanças sinápticas duradouras - ou LTP. (A descrição fornecida aqui foi simplificada. A LTP é hoje tema de amplas investigações.)
Como a estimulação tetânica aplicada por Bliss e seus colegas não ocorria naturalmente no cérebro, restava a questão de saber como a LTP era obtida em condições normais. Em 1986, John Larson e Gary S. Lynch, da University of California, em Irvine, e Gregory Rose e Thomas V Dunwiddie, da University of Colorado, em Denver, sugeriram que a ocorrência de LTP no hipocampo estava ligada ao ritmo teta. Eles aplicaram um pequeno número de estímulos elétricos às células de CAI do hipocampo de um rato e produziram LTP, mas somente quando os estímulos eram separados pelo lapso de tempo normal entre duas ondas teta - aproximadamente 200 milésimos de segundo. O ritmo teta e, aparentemente, o meio natural pelo qual o receptor NMDA é ativado em neurônios no hipocampo.
Pesquisas feitas em meu laboratório da Rockefeller University reproduziram as descobertas de Larson e Lynch, mas desta vez nas células granulares do hipocampo. Constantine Pavlides, Yoram J. Greenstein e eu demonstramos que a LTP dependia da presença e da fase do ritmo teta. Se fossem aplicados estímulos elétricos às células no pico da onda teta, LTP era induzida. Se o mesmo estímulo fosse aplicado no ponto mais baixo das ondas - ou na ausência de ritmo teta - não se induzia LTP.
Surgia, assim, um quadro coerente sobre o processamento da memória. Por exemplo, quando um rato explorando os neurônios do tronco encefálico ativam o ritmo teta. Os sinais de entrada olfativos (que no rato estão sincronizados com o ritmo teta, assim como o movimento das vibrissas) e outras informações sensoriais convergem ao córtex entorrinal e ao hipocampo. Elas são aí divididas em "bites" de 200 milésimos de segundo pelo ritmo teta. Os receptores NMDA, agindo com o ritmo teta, permitem a armazenagem de longo prazo dessa informação.
Um processo similar ocorre durante o sono REM. Embora não haja entrada de informação ou movimento durante o sono REM, a rede natural do neocórtex e do hipocampo sofre mais uma vez a ação marca-passo do ritmo teta. O ritmo teta pode produzir mudanças duradouras na memória.
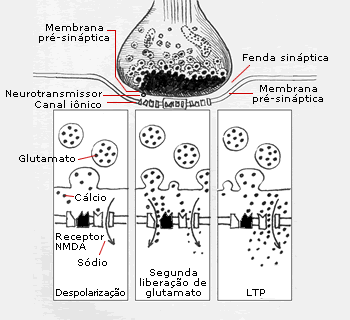
A ativação do receptor NMDA induz a potencialização a longo prazo (LTP), um modelo para a memória. A liberação do neurotransmissor glutamato (quadro à esquerda) abre um canal associado a um receptor não-NMDA (N-metil-D-aspartato), permitindo o influxo de sódio, que despolariza o neurônio. Se uma nova liberação de glutamato ocorrer enquanto a célula estiver despolarizada (quadro central), o receptor NMDA abre um segundo canal, que permite o influxo de cálcio e leva à LTP. A LTP é resultado do aumento do influxo de sódio através do canal associado a um receptor não-NMDA (quador à direita) e do subseqüente aumento da despolarização da célula.